Integumentary (surface of the bird)
The Integumentary System
The integumentary system consists of the skin, the feathers and the appendages (claws and beak). The skin covers the majority of the body and contains glands in the outer ear canal and the preen gland at the base of the tail, that the bird uses to preen its feathers. The integumentary system is very important in providing protection to the bird from a number of potentially dangerous situations. The functions provided by the integumentary system include:
- A barrier between the external environment and the internal systems and organs thus provides support and protection from infection by microorganisms and from physical injury.
- Excellent thermal insulation to help regulate body temperature in a variable environment.
- Numerous nerve endings for the senses to enable the bird to be aware of potentially harmful situations.
- Pigments for display and protection from the elements.
- The compounds capable of conversion into vitamin D when exposed to sunlight.
The skin
The skin is composed of three tissues or layers of cells:
- The epidermis – the outermost tissue itself consisting of three separate layers of cells.
- The dermis or corium – the middle tissue.
- The hypodermis – the innermost layer or tissue.
The fowl’s skin is divided into a number of separate areas where the skin has been modified to some extent to be able to carry out special functions. These areas are:
- The feathered skin.
- The scale covered skin on the lower legs and feet.
- The hard, horny areas of the beak and toenails.
- The pad of the foot (or plantar).
- The skin of the comb and wattles.
The feathered skin is also divided into a number of special areas or tracts – those where feathers actually do grow and other areas where they do not grow but which appear as such because they are covered by feathers. The areas where they do grow are called pterylae and the areas where they do not grow are called apteria.
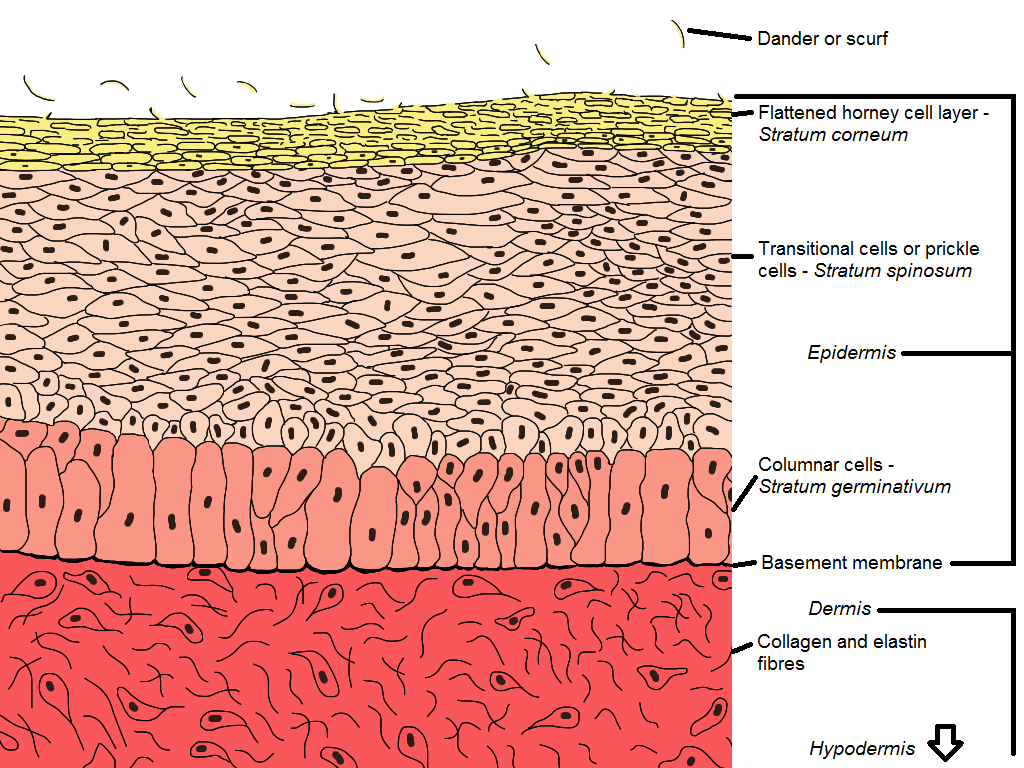
Cross-section of bird skin
Structure
The skin is mainly composed of two different tissues:
- Connective tissue of the dermis and hypodermis where the cells are widely spaced.
- Cellular stratified epithelium of the epidermis, which make up the horny outer layer of the skin and forms the feathers.
The different skin types contain different amounts and distribution of special compounds mainly collagen, elastin and keratin.
The epidermis consists of three separate layers:
- The flattened, horny cell layer – the outermost layer.
- The transitional layer – joins the outer layer to the inner layer. It is here that the cells formed in the third innermost layer are transformed into the hard, horny type found in the outermost layer.
- The columnar cell layer – this is the innermost layer of the epidermis and is the germinative layer where the cells of the epidermis are formed.
Dermis
The dermis is relatively thin and shows a uniform, microscopic structure. This is different to many other animals that have both densely packed and loose layers of cells. The main component of the dermis is collagen with a small amount of elastin. The hypodermis contains fewer cells and is more loosely arranged than the dermis. Fat is stored here in special adipose cells (adipose tissue is fat tissue). Air spaces found in the hypodermis connect with air sacs of the respiratory system to enhance the ability to fly.
Skin surface cells are replaced continuously through life by mitosis in the epidermal germinative layer. As an outer or surface layer wears away or is lost, the cell layer beneath replaces it. The tissue lost in this way forms part of the scurf or dander that provides nutrition for many parasites such as lice and a place for the survival of disease causing viruses such as Marek’s Disease. Almost all of the activity of the epidermis is devoted to keratinisation that involves two processes:
- The synthesis or manufacture of the very durable fibrous protein, keratin that gives the skin surface its strength and resistance to normal wear and tear.
- The death of the cell – the final stages of keratinisation occurs in the transitional layer of the epidermis. As the cells reach the stage where they become part of the horny cell layer they die.
The skin has many nerve endings located in it. These are mainly located in the outer layers of the dermis and provide good protection from potentially harmful situations by making the bird aware of their presence. The dermis is also quite well supplied with blood vessels that do not reach into the epidermis. Nutrients, respiratory gases, hormones and other compounds pass by diffusion into the live inner cells of the epidermis.
Skin of the feathered areas
The skin is usually covered by the feathers is therefore normally protected and hence is thinner. Over the wings and thighs, the skin is more closely joined to underlying tissue than over the rest of the body. The epidermis is about 12 cells thick with the horny outer layer being about 5 cells, the transitional layer being about two and the inner, germinative layer being about 4-6 cells.
Skin of the legs and feet
The lower legs, including the upper sides of the toes, are covered by skin carrying scales attached to the epidermis. In those breeds with feathered legs the feathers grow out of the margins of the scales. The scales are considered to be a carry over from the bird’s reptilian ancestors. Scales are very rich in keratin.
The skin of the foot pad or planter
The skin beneath the toes and the foot is specialised to withstand compression caused by the weight of the bird when standing and abrasion caused when the foot comes in contact with rough surfaces as the bird moves about. The skin of this area is quite thick but retains a high degree of flexibility.
The skin of the beak
The beak is composed of two parts:
- The internal skeleton of bone
- A closely attached layer of highly keratinised skin over the bony skeleton
The epidermis of the beak develops a thick, horny layer (called rhampotheca) made of sheets of flattened cells firmly attached to each other. The strength of the beak keratin is partly due to its special make-up. The hard egg tooth that a hatching chicken uses to break out of the shell is made of strongly calcified keratin cells. It is not a true tooth.
The skin of the claws, wattles and comb
The claws are keratinised epidermal appendages of the digits or toes. The claw root epidermis gives rise to columns of flattened keratinised cells that remain firmly stuck together.These are accessory sexual epidermal appendages on the head and neck. They develop as a result of the production of hormones by the sex organs of the male and female when these are functional. The very rich supply of blood vessels of these appendages is the cause of the bright red colour. In very high temperatures the bird dips these organs in the water supply as an aid to cooling.
The uropygial or preen gland
This is a two-lobed, pea-sized structure that develops from the epidermis at the base of the tail above the pygostyle or last vertebra. It produces and discharges a fatty secretion through a duct opening on a small papilla or pimple located on the surface of the skin. It is one of the only two groups of glands found in the skin of the fowl as far as is known (the other group is located in the skin of the outer ear canal). The secretion of this gland is holocrine and is made from disintegrated cells. As new cells are formed they are pushed towards the centre of the gland where they create the gland secretion that contains:
- Saponifiable lipids (special fats that can be broken down by an alkali)
- Non-saponifiable lipids
- Lecithin (special phosphorus based compounds)
- Cell degradation or breakdown products
The secretion is similar to that of mammalian sebaceous glands and appears to carry out a similar function. It is collected by the beak during feather preening and is used to oil the feathers. This is very important to aquatic birds for water proofing but not so to terrestrial birds.
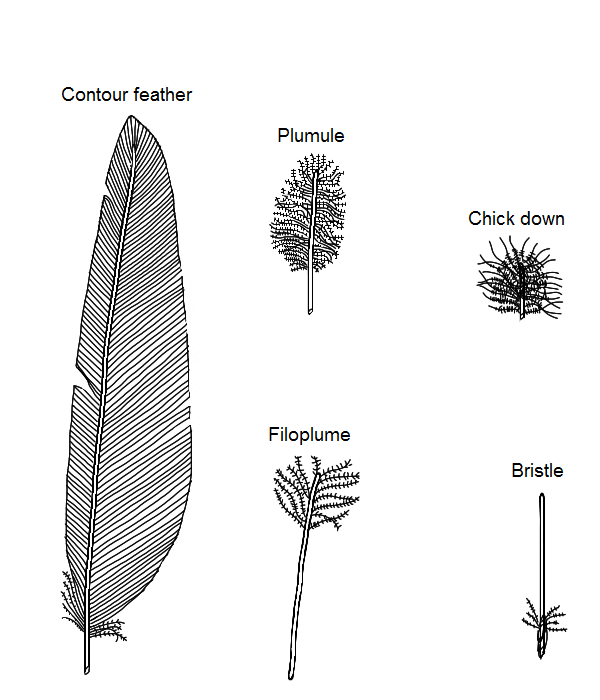
Types of chicken feathers
Feathers
Feathers are confined to precise tracts or areas called pterylae. Areas devoid of feathers are called apteria. Smooth muscles attached to the feather follicle from which the feather emerges cause erection of the feather during cold periods thus increasing the volume of air trapped therein. This in turn increases the insulation capacity of the feather layer. This ability to control the feathers is also utilised in periods of hot weather when the birds release hot air from among the feathers by erecting them.
In fowls there are five types of feathers:
- Contour feathers
- Plumules
- Filoplumes
- Down (chick)
- Bristles
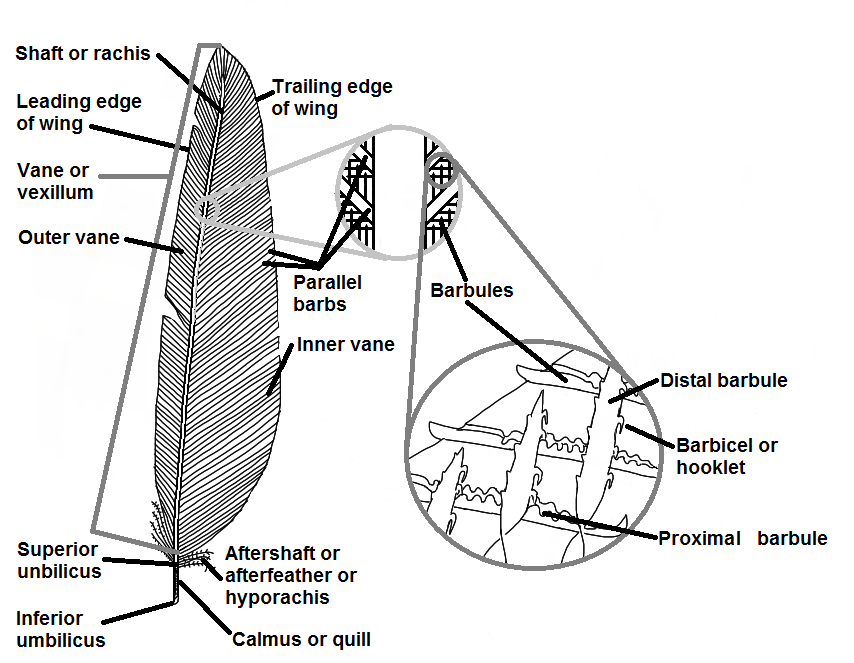
Parts of contour feather
Contour feathers
These consist of:
- The shaft or rachis.
- The outer and inner vanes, or vexillum, which slope obliquely towards the tip and on which are:
- Parallel barbs.
- Barbules.
- Barbicels or hooklets that hold the barbs together to form the vane. Each barb carries two rows of barbules. The distal row (those growing from that side of the barb facing towards the tip of the feather) has microscopic hooklets that hook onto the proximal barbules of the next barb. These hooklets are called barbicels. This interlocking gives a continuous, strong, smooth surface.
- The calamus or quill (lower shaft) which is rounded, transparent, hollow and with a series of conical scales in its interior.
- Inferior umbilicus – forms as a papilla of epidermis during the feather formation and which, in time, disappears and leaves a hole at the proximal end of the feather.
- Superior umbilicus – a hole in the shaft at the junction of the shaft and the vanes.
- The aftershaft or hyporachis – a small feather-like tuft located near the superior umbilicus.
- Rachis – a four sided, solid, tapering and elastic shaft running the length of the vane. A longitudinal groove facing towards the body when the feathers are in the normal position runs the length of the rachis.
All contour feathers are not the same. The main wing feathers, called remiges, have the posterior portion of the vane broader than the anterior portion (anterior = front; posterior = rear or back). They are divided into the primary and secondary flight feathers separated by the smaller axial feather. The primary flight feathers are located on the trailing edge of the outer part of the wing and the secondaries are similarly located but closest to the body. There are 10 primary feathers on each wing of the normal adult fowl. The large feathers of the tail are called reticles. With these feathers the vanes are of equal width. Special feathers called coverts cover the bases of the remiges and rectrices e.g. wing coverts and tail coverts.
Plumules
Plumules are found beneath the contour feathers where they form a soft, downy undercoat. They have a short shaft with radiating, free barbs and barbules. They have no interlocking hooklets or barbicels. These feathers provide the depth to the coat and play a large part in trapping the air and holding it still, and thus improve their ability to conserve warmth.
Filoplumes
The filoplumes are very small and hair-like with rudimentary barbs and barbules which are confined to the apex. These feathers are often quite troublesome during the processing of live poultry into poultry meat as they are very difficult to remove. If they are dark in colour they are easy to see and tend to detract from the appearance of the carcass.
Chick down
When a chicken hatches it has a coat of short, fluffy down which closely resembles plumules in that it has multiple radiating barbs. At first moult the juvenile feather emerges from the same follicle to replace the fluff. The outermost juvenile feathers resemble adult contour feathers but with a softer texture. The order of emergence of juvenile feathers varies with different pterylae and is related to when the feather follicles appear in these areas during embryonic development. Juvenile plumage is eventually replaced by definitive (final) plumage. The number of moults before adult contour feathers are formed is specific for each follicle, but varies between pterylae.
Bristles
Bristles are found around the mouth and eyes of chickens and are thought to be primarily used to aid the sensory ability of birds and protect sensitive areas. They are comparable to mammalian eylashes.
The feather follicle
Feathers are keratinised epidermal derivatives related to epidermal scales. Each follicle develops in the embryo as a dome-shaped projection of the epidermis over a raised dermal papilla or pimple. The dome later sinks into the skin by a downward growth of its rim so that eventually it lies in a short tube beneath the skin surface. The base of the rim, called the collar, continues to show rapid cell division during feather formation and constitutes the germinal region of the follicle. The cells that form the feather by differential growth become arranged into structural patterns of calamus and rachis, barbs, barbules and barbicels.
During the development phase of the feather, the dermis within the follicle is highly vascularised (many blood vessels to supply the nutrients required for feather formation). But after the feather’s cells become keratinised, this part of the dermis (in the follicle) dries up and forms the pulp found inside the calamus. Finally, the feather sheath (formed by the horny layer of the epidermis) splits and the new feather unfurls. No new follicles develop after hatching.
Colouration of feathers
Black, brown and reddish brown colours are due to the granular pigment, melanin, deposited in the keratinised feather cells. Melanin is formed from tyrosine (a non-essential amino acid) by special pigment cells called melanocytes. These cells enter the dermis during embryonic development and congregate beneath the feather follicles. Melanin granules move into epidermal cells prior to keratinisation. The melanin colour may be black or reddish (called phaemelanin) – the chemical differences between the two are unknown.
Carotenoids
Carotenoids are the other important skin pigments. These are yellow to orange lipid pigments contained in free fats and are found in some dietary ingredients. Carotenoids in the skin, beak and legs are responsible for the yellow colour of these tissues and organs and are also responsible for the yellow colour of egg yolk. Carotenoids are very commonly found in many plants such as maize, carrots and lucerne leaf.
Colour patterns
Colour patterns are largely dependent upon the distribution of melanin and other pigments. Colour bands of light and dark as seen in barred plymouth rocks are due to cyclic changes in melanin deposition during feather growth. The colour patterns of some breeds such as brown leghorns, show sex dimorphism (the colour depends on the sex). Genes are also responsible for mutant feather types such as frizzle and woolly feather types. Colours and colour patterns are controlled by two groups of genes:
- Those controlling melanin formation
- Those controlling the patterns in which the colours are deposited
The physiology of moulting
Each feather follicle undergoes cyclical changes of growth called anagen and rest period, called telogen. The growth phase occurs prior to the moult of old plumage. Once growth has occurred the completed feather remains attached to the base of the resting follicle by its quill or calamus.
Telogen period
At the end of the telogen period, the germinal cells again enter an anagen phase. As the new feather develops the quill of the old is pushed out of the follicle canal but remains attached to the horny sheath of the new feather. When the sheath splits to reveal the new feather the old inevitably falls away (is moulted). Thus the moulting of feathers is a mechanical process dependent upon the growth of a new feather. Completed plumage is always in the telogen phase. Plucking feathers from the bird can induce the anagen phase.
Feather growth
Sex-linked genes control when feather growth is initiated and down replaced. Chickens are described as being fast or slow feathering or, more correctly, as early or late feathering. There is some form of local control in the moulting of feathers in close proximity, such as a hormone. In wild birds there is a growth centre in each pterylum (pterylae) that provides an ordered sequence to the normal moult. This control is not so evident in fowls although the proximal flight feathers moult first.
There is also a degree of control between pterylae. Bilaterally symmetrical pterylae have synchronised moulting which is an important feature in flying birds. The sequence of pterylae starts with the wing feathers and spreads to the body and ends at the neck and head. In fowls, the moult is a seasonal occurrence in autumn. However, environmental conditions may initiate a moult at other times. Fowls in good production are prone to a moult when stress is applied. Males moult before females.
Thyroxine
Thyroxine, the hormone of the thyroid gland plays an important part in the growth, differentiation and patterning of feathers by increasing the metabolic activity of the feather forming cells. Chicken down is not influenced by thyroxine and only the feathers of juveniles and adults are dependent on having adequate levels available. Wing feather growth is not dependent and, after the removal of the thyroid gland, all feather growth in adults ceases except on the wings. Giving thyroxine induces a moult by speeding up the anagen phase with females being more sensitive than males.
Oestrogen
High levels of oestrogen (female sex hormone), such as what occurs during egg laying, retard feather formation and maximum egg production occurs when the follicles are in the telogen phase. A reduction in production is usually associated with the anagen phase. Oestrogen and testosterone influence feather development and are responsible for the differences that occur between males and females. In the absence of androgens, males develop “female” type feathers. Removal of the gonads will produce a situation where the feathers are being replaced continuously rather than on an annual basis i.e. an annual moult.
Light pattern
The light pattern to which the birds are subjected also affects feather development and moulting because of its effect on the pituitary gland. This, in turn, controls the production of gonadotropins by the ovaries and testes, and thyroxine by the thyroid gland. Increasing day length stimulates the production of the gonadotropins and thyroxine while decreasing day length causes a reduction in their production.
You May Also Like

The Poultry Hub Australia profoundly acknowledges and respects that its foundations, both people and facilities, are established on land rich in the history and traditions of the world’s oldest living culture. PHA values and respects Indigenous knowledge, understanding its importance in our shared history. We acknowledge the strength, resilience, and contributions of the Aboriginal community, we pay our tributes to the Aboriginal Elders – those who guided us in the past, those who lead us today, and those who will enlighten our paths in the future.