Skeletal System
Skeletal System
The skeletal system provides the strong framework for the support and protection of the remainder of the systems, organs and tissues that make up the body of the fowl. Bird bones that are homologous to bones found in other animals have evolved over time to enhance the ability of the bird to fly. While fowls are not able to fly well, they still retain that ability to some extent. These modifications include:
- Pneumatic bones where the air sacs of the respiratory system connect with the cavity of some of the long bones.
- Fusion of some vertebral sections to provide the rigidity required for flight.
- The sternum provides a large surface area for the strong attachment of the main muscles used for flight.
- Compared to other animals, the size of the head has been reduced significantly as a large head would be a hindrance when flying.
- The neck is quite long in most bird species to enable:
- Protection of the delicate tissues of the brain from too much jarring when landing. The flexibility of the neck acts as a shock absorber.
- The bird to reach food located on the ground – a rigid body makes this simple activity more difficult.
- The bird to adjust its centre of gravity when the bird changes from the upright position of walking or perching to the more horizontal position of flight.
- The long tail of many other animals has been reduced to a very short section of fused bones called the pygostyle.
- The ribs have been modified by the inclusion of the uncinate process – a rearward projection of bone – which gives strength to the rib cage.
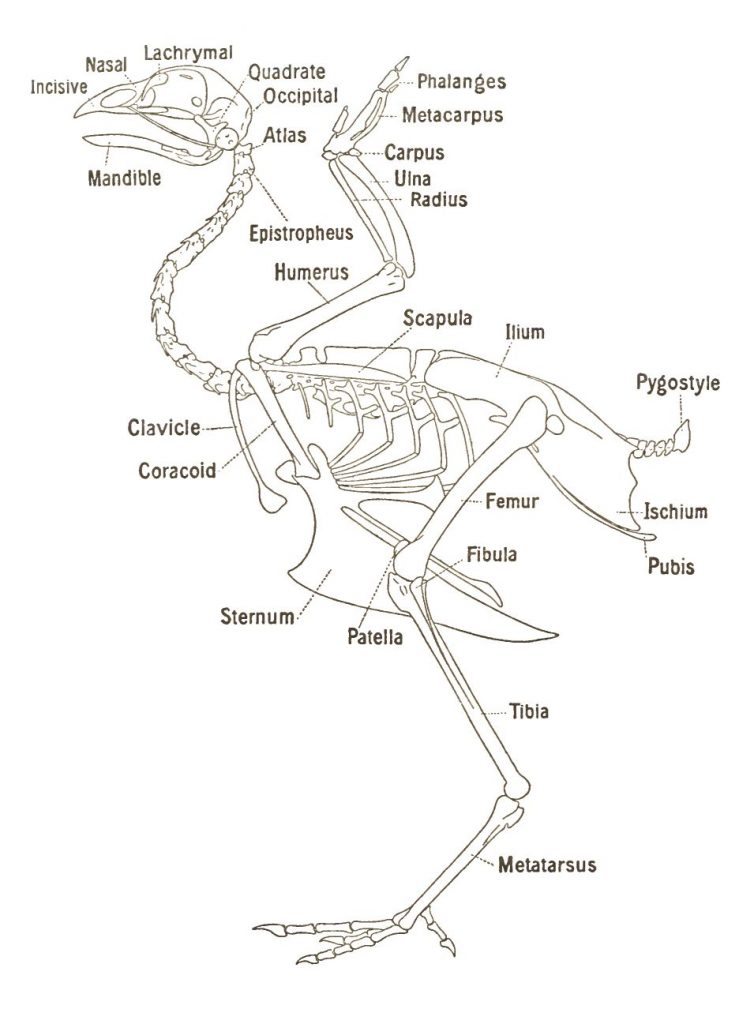
Fowl skeleton
Vertebrae
The vertebral column has 39 separate bones and is divided into five sections or groups – the cervical vertebrae, the thoracic vertebrae, the lumbar vertebrae, the sacral vertebrae and the coccygeal vertebrae. The vertebral column is often described by way of the vertebral formula that is:
C13, T7, L+S14 & C5 = 39 (The vertebral formula refers to the number of vertebrae in each of the major sections of the column)
Atlas
The atlas-axis at the base of the skull is quite different to all other bones of the vertebral column. The atlas is small and ring-like with a deep cavity for articulation (moving together) with a single projection, or condyle (a rounded projection found on many bones), at the base of the skull.
Axis
The axis, or epistropheus, is also specialised. It is short and projects from the cranial end and passes through the ring-like atlas. It possess a small process, the dens, which allows the axis to articulate with the occipital condyle. This joint allows the head to turn on the neck.
Thoracic and Lumbar
The seven thoracic vertebrae carry the ribs and all except the last have large ventral processes for the attachment of muscles. Fusion of the second to fifth vertebrae provides rigidity for the structural strength necessary for flight. It is very difficult to separate the seventh thoracic, lumber, sacral and first coccygeal vertebrae because they are so closely fused for strength. Consequently, the lumbar and sacral vertebrae are usually treated as one group. The last of the coccygeal are fused to form the pygostyle and provides a strong foundation for the tail feathers.
Ribs
There are seven pairs of ribs originating on the thoracic vertebrae and all but the first, second (and sometimes the seventh) do not reach the sternum, which is their other attachment point. The third to the sixth have two segments:
- Vertebral segment
- Sternal segment
All except the first and last have uncinate (meaning hooked or bent)processes that project backward over the outer surface of the next rib and connect to it by a ligament, which adds strength to the thoracic cavity.
Sternum
The sternum or breastbone is a complex shape and has been described as a quadrilateral, curved plate with processes projecting from each angle and from the middle of the cranial and caudal border.
Metasternum
The caudal medial projection, or metasternum, is the longest projection and carries the tall plate like ridge or sternal crest that runs from front to back on its ventral surface. This crest is more commonly called the keel bone and provides a suitable attachment for the major muscles of flight – the pectoralis and subcontractors muscles.
Skull
The skull is divided into two regions:
- Rounded cranium
- Conical facial region
The distinction between the two is very easy because of the two large orbits or openings into which the eyes fit. Two very thin bones, the sphenoid and ethmoid bones, together form the very thin septum that separates these orbits.
Brain capacity
The cranial external appearance suggests a larger brain capacity than there really is. This is because the bone is formed by two layers of dense bone separated by a layer of very spongy bone. The spongy bone contains pockets of air derived from the eustachian tubes that connect the upper respiratory system with the middle ear.
Eardrum, spinal cord, carotid arteries and jugular veins
Large hemispherical cavities at the rear called the tympanic cavities form the location for the eardrum. Openings at the base of the cranium provide for the direct connection of the brain with the spinal cord, and common openings in each lateral part of the occipital bone provide for the carotid arteries and jugular veins.
Eyes and tongue
The two optic nerves enter the cranial cavity by one common opening. The hyoid bone is found beneath the skull and forms the framework for the tongue. It has three major sections – the entoglossal, the basi-hyal and the third section is itself in three parts. The entoglossal is contained within the tongue and a movable joint connects it with the basi-hyal and hence the remainder of the bone.
The forelimb or wing
The skeleton of both the forelimb or wing and the leg is very similar to those of theropod dinosaurs. They are essentially pentadactyl (five digits) limbs modified for the special purpose of flight and specialised feeding.
The wing skeleton may be divided into:
- The shoulder girdle:
- Scapula
- Coracoid
- Clavicle
- The wing:
- The upper arm – or humerus
- The forearm – radius and ulna
- The manus or hand – carpus, metacarpus and digits
Scapula and coracoid
The scapula is narrow, thin and slightly curved which is unlike the shoulder blade of other animals. At the cranial end a section of a cavity receives the head of the humerus. The coracoid is the strongest bone of the shoulder girdle. One end carries a flattened articular surface to fit into the sternum. At this end is a hole, or foramen, for the connection of the clavicular air sac (a pneumatic bone). The other end connects with the clavicle.
Clavicle
The clavicle or collarbone is thin, rod-like and slightly bent. Its upper, or dorsal, end is connected with the coracoid bone. The other end is joined to that of the other wing to form the “wish-bone”. The combined clavicles form a bone called the furcula that is capable of acting like a spring and provides a firm base of support for the wing. An opening formed by the shape of the scapula, coracoid and clavicle where they join provides a passage for the tendon of the muscle supracoracoides for connection to the humerus.
Humerus, radius and ulna
The humerus is a large long bone with an ovoid head for articulation with the scapula, coracoid and clavicle. It is a pneumatic bone with a connection with the clavicular air sac. The two bones of the forearm are the ulna (the thicker and longer), and the radius that lies laterally to the ulna. The large space between the ulna and radius is called the interosseus space.
Manus (hand)
The manus, or hand, consists of the carpus, metacarpus and the digits. The carpus of an adult contains only two bones – the ulnar and radial (note different spelling) that represent the proximal row of mammalian carpal bones. In the embryo, cartilaginous nodules represent the distal row but these fuse with the metacarpus and disappear. In the adult, the metacarpus is in the form of a single bone that is produced by the union of three elements that correspond to the first, second and third metacarpal bones of the theropod limb. Modification and fusion has reduced the number of recognisable digits to three which are carried by the metacarpals.
The leg
The significant features of the skeleton of this limb are:
-
- The hipbone is firmly fixed to the vertebral column.
- There is no ventral union between the two hipbones.
- There is no independent tarsus.
The leg skeleton may be divided into:
- The pelvic girdle or hip bones:
- Ileum
- Ischium
- Pubis or pin bones
- The leg:
- Femur or thighbone
- Tibia and fibia
- Pes or foot – tarsus, metatarsus and digits or toes
Unlike with other animals, such as mammals, the bones of the opposing sides of the pelvis do not meet on a mid ventral line. They are at their widest separation in this location. Hence the term “girdle” could be said to be a misnomer. This difference could be seen as a weakness, however it is compensated for by the extensive fusion of the hipbone and the vertebral column. The hipbone, in fact, consists of three bones – the ileum, the ischium and the pubis which all meet at a deep concavity called the acetabulum, into which the head of the femur fits.
Ileum, ischium and pubis
The ileum is fused to the last thoracic, lumbar and sacral vertebrae to provide strength and rigidity. The ischium is much smaller and is continuous with the ileum. An opening provides access for the sciatic nerve. The pubis is a narrow strip of bone that runs along the border of the ischeum to which it is joined for a short distance only. The free posterior end projects backward slightly beyond the ischeum to form the pin or pubic bones.
Femur, tibia and fibula
The femur is a typical long bone and is quite thick and bent. The proximal end has a prominent head that fits loosely into the acetabulum. The distal end carries the deep pulley shaped surface for the patella (knee cap) and two convex condyles that articulate with the bones of the lower leg. The tibia is a much longer bone than the fibula and is much thicker at the proximal end than it is at the distal end. The proximal row of tarsal bones is fused to the distal end of the tibia. The fibula is greatly reduced compared to other animals and consists of a slender spicule (needle-like) and has a flattened head for attachment to the proximal head of the tibia.
Feet
In the adult fowl there is no independent tarsus. In the embryo, the two rows exist but the proximal fuses with the tibia. The adult metatarsus is a long bone actually formed by the union of the second, third and fourth metatarsal bones. In male chickens a small conical projection on the medial side supports the spur. Most fowl breeds have four digits – three facing forwards and one facing back. There are some breeds that have five digits.
Bone formation
The skeleton provides support and protection for the remainder of the systems and tissue. Bone is living tissue and its structure is largely affected by the nature of stresses placed upon it. The chemical composition is also quite variable although it mainly consists of calcium and phosphorus in the form of hydroxyapetite [3Ca3(PO4)2.Ca(OH)2] crystals deposited on a fine matrix of collagen fibres, along with other cell types.
Deposition and adsorption of bone
The metabolic activity in bones is continuous and the microscopic structure is constantly changing. Small cells, called osteoblasts, are responsible for depositing new bone tissue, while large polynucleate cells, called osteoclasts, resorb existing bone. Other cells, called osteocytes, found in the calcified mass of bone, help maintain the bone structure. Thus the skeleton is a major reservoir of calcium and phosphorus. Therefore, it is very important to maintain the proper levels of these minerals in the diet (99% of body calcium and 80% of body phosphorus are stored in the skeleton). Sodium and magnesium are other minerals of importance in bone structure. These may be drawn upon when the diet of the animal contains inadequate supplies.
Microstructure
The microstructure of the bone changes continuously as bone is a target for a number of influences. Chemicals, in particular calcium and phosphorus, are continuously added or removed from the bone. Other influences include:
-
- The level of certain hormones e.g. growth hormone, parathyroid hormone, calcitonin, oestrogenic and androgenic hormones in the blood
- The level of vitamin D in the diet
Young chickens are very sensitive to vitamin D deficiency. This vitamin is required by the chicken for the assimilation and use of calcium and any deficiency will be seen as a typical calcium deficiency, such as rickets. Vitamin D is found in a number of slightly different forms and cholecalciferol (D3) is ten times more active than ergocalciferol (D2) in preventing rickets.
Ergosterol, a compound under the skin of animals including poultry, is converted to usable Vitamin D by the rays of the sun. In the layer hen, the skeleton is particularly vulnerable to the demands made for calcium for eggshell formation.
Stages of bone development
All bones of the body are formed in pre-existing tissues that they either replace or use in their structure. It is usual to find that bones pass through three stages as they develop:
-
- Prechondral or membranous stage
- Chondral or cartilaginous stage
- Ossification stage (bone formation)
Most of the bones of the fowl go through the cartilaginous stage. A few such as the bones of the skull omit this stage. As far as the chicken is concerned, the membranous stage takes place in the egg during embryo development. Only the cartilaginous stage and the ossification stages are easily identified.
The secretion of special cells called chondroblasts, brings about cartilage formation. The ossification process then hardens the cartilage when the bone takes up minerals, mainly calcium carbonate. Long bones increase in length by the ossification process. In birds much of the bone is laid down in successive layers to form dense, compact bone covered by the cellular periosteum on the outside. Long bones are usually hollow with the hollow filled with bone marrow and extensions of the air sacs.
Compact bone is modified by the formation of special cavities that eventually mineralise by depositing concentric layers of new bone. The new structure is called the haversian system. If a transverse section (slice) of bone is examined under a microscope, a large number of small canals that run more or less parallel to the long axis will be seen. These canals are called haversian canals and carry the blood vessels and nerve fibres. Surrounding these canals are plates of bone and between the plates are small spaces called lacunae. A special bone cell called an osteocyte is found in each lacunae. Nutrients pass from the blood vessels in the haversian canals to the bone through small canals called canaliculi.
Medullary bone (layer fatigue)
A very unique feature of the female avian skeleton is the way the bird lays down a supply of highly unstable secondary bone in the marrow cavities of bones during the reproductive period. This bone is called medullary bone and because of its instability, provides a very ready source of calcium for eggshells. In a production hen, not enough calcium can be absorbed across the intestinal wall in one day supply to satisfy the requirements for production of an egg shell. Without this medullary bone the eggshells would be very thin and weak.
Medullary bone starts to develop about 10-14 days before the first egg is laid as a result of the presence of oestrogen and androgen in the blood as the pullet reaches sexual maturity. This function remains for the length of her laying life. Approximately two weeks before egg production starts the pullet flock should be changed from the growing to the layer diet which is higher in calcium. If pullets come into production at too young an age, they may deplete body reserves of calcium that may result in thinner shelled eggs and /or a drop in production. This should not occur if production is delayed to an age best suited to the strain of layer.
If a calcium deficient diet is provided to a layer, it will deplete her skeleton of calcium and thus make the bird significantly weaker. Eggshells will also become thinner and production will decline until it ceases altogether. A condition of paralysis, called cage layer fatigue, may be seen in layers housed in laying cages. When seen, it is always associated with birds in very high production and takes the form of muscular paralysis and osteoporosis (weak bones). While the cause is not fully understood, the birds usually recover quite quickly when taken out of the cages and placed on the floor for a short period. The weaker shells of eggs from older hens are caused by a loss in efficiency by the oviduct shell glands in their production of the shells.
References
-
- Bradley, OC (1915) The Structure of the Fowl, A & C Black, London, UK.
- Storer, TI, Stebbins, RC, Nybakken, JW and Usinger, RL (1976) Elements of Zoology, 4th Ed, McGraw Hill, New York, USA.
- Wilson, BW (ed) (1980) Birds, Freeman, San Francisco, USA.

The Poultry Hub Australia profoundly acknowledges and respects that its foundations, both people and facilities, are established on land rich in the history and traditions of the world’s oldest living culture. PHA values and respects Indigenous knowledge, understanding its importance in our shared history. We acknowledge the strength, resilience, and contributions of the Aboriginal community, we pay our tributes to the Aboriginal Elders – those who guided us in the past, those who lead us today, and those who will enlighten our paths in the future.